
Cargo recognition and cargo-mediated regulation of unconventional myosins
2014.09.17Lu, Q., Li, J., & Zhang, M. (2014). Accounts of chemical research, 47(10), 3061-3070.
Organized motions are hallmarks of living organisms. Such motions range from collective cell movements during development and muscle contractions at the macroscopic scale all the way down to cellular cargo (e.g., various biomolecules and organelles) transportation and mechanoforce sensing at more microscopic scales. Energy required for these biological motions is almost invariably provided by cellular chemical fuels in the form of nucleotide triphosphate. Biological systems have designed a group of nanoscale engines, known as molecular motors, to convert cellular chemical fuels into mechanical energy. Molecular motors come in various forms including cytoskeleton motors (myosin, kinesin, and dynein), nucleic-acid-based motors, cellular membrane-based rotary motors, and so on. The main focus of this Account is one subfamily of actin filament-based motors called unconventional myosins (other than muscle myosin II, the remaining myosins are collectively referred to as unconventional myosins). In general, myosins can use ATP to fuel two types of mechanomotions: dynamic tethering actin filaments with various cellular compartments or structures and actin filament-based intracellular transport.
In contrast to rich knowledge accumulated over many decades on ATP hydrolyzing motor heads and their interactions with actin filaments, how various myosins recognize their specific cargoes and whether and how cargoes can in return regulate functions of motors are less understood. Nonetheless, a series of biochemical and structural investigations in the past few years, including works from our own laboratory, begin to shed lights on these latter questions.
Some myosins (e.g., myosin-VI) can function both as cellular transporters and as mechanical tethers. To function as a processive transporter, myosins need to form dimers or multimers. To be a mechanical tether, a monomeric myosin is sufficient. It has been shown for myosin-VI that its cellular cargo proteins can play critical roles in determining the motor properties. Dab2, an adaptor protein linking endocytic vesicles with actin-filament-bound myosin-VI, can induce the motor to form a transport competent dimer. Such a cargo-mediated dimerization mechanism has also been observed in other myosins including myosin-V and myosin-VIIa.
The tail domains of myosins are very diverse both in their lengths and protein domain compositions and thus enable motors to engage a broad range of different cellular cargoes. Remarkably, the cargo binding tail of one myosin alone often can bind to multiple distinct target proteins. A series of atomic structures of myosin-V/cargo complexes solved recently reveals that the globular cargo binding tail of the motor contains a number of nonoverlapping target recognition sites for binding to its cargoes including melanophilin, vesicle adaptors RILPL2, and vesicle-bound GTPase Rab11. The structures of the MyTH4–FERM tandems from myosin-VIIa and myosin-X in complex with their respective targets reveal that MyTH4 and FERM domains extensively interact with each other forming structural and functional supramodules in both motors and demonstrate that the structurally similar MyTH4–FERM tandems of the two motors display totally different target binding modes. These structural studies have also shed light on why numerous mutations found in these myosins can cause devastating human diseases such as deafness and blindness, intellectual disabilities, immune disorders, and diabetes.
- Recommend
-
2025-10-22
IQSEC2/BRAG1 may modulate postsynaptic density assembly through Ca2+-induced phase separation.
-
2025-08-22
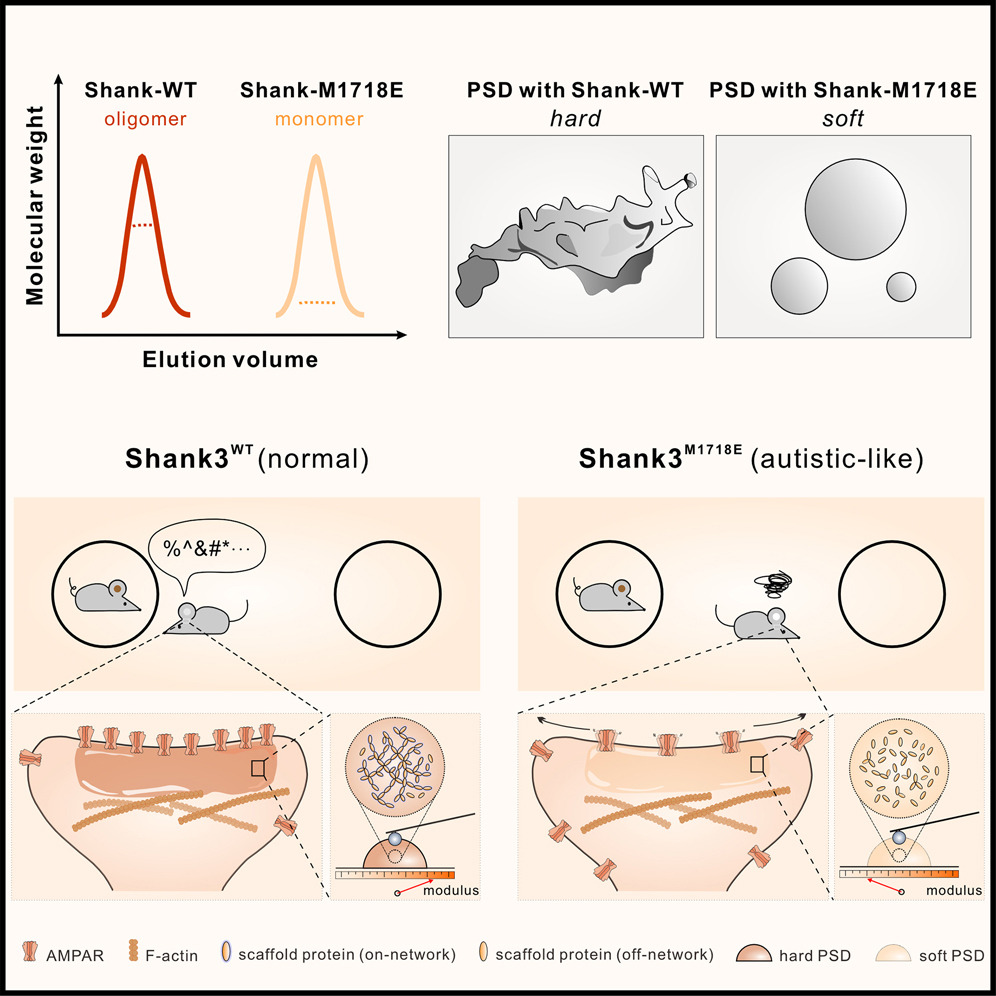
Shank3 oligomerization governs material properties of the postsynaptic density condensate and synaptic plasticity.
-
2025-08-21
Modulating synaptic glutamate receptors by targeting network nodes of the postsynaptic density condensate.
-
2025-08-19
Current practices in the study of biomolecular condensates: a community comment.
-
2025-06-10
Phase separation instead of binding strength determines target specificities of MAGUKs.